Animateurs

Florian Frugier
IPS2
Directeur de Recherche au CNRS, est responsable d’une équipe travaillant à l’Institut des Sciences des Plantes de Paris-Saclay (IPS2, Université Paris-Saclay) sur les voies de signalisation végétales agissant localement et à longue distance (systémiquement) pour réguler la nodulation symbiotique chez les légumineuses, en relation avec la disponibilité en azote.

Sandra Bensmihen
LIPME
Chercheuse au LIPME, Toulouse. Elle cherche à comprendre comment les signaux symbiotiques produits par les rhizobia et les champignons mycorhiziens peuvent stimuler le développement racinaire des plantes (légumineuses ou non) et si ces mécanismes sont proches de ceux qui contrôlent l’organogenèse du nodule et la mise en place de ces symbioses.
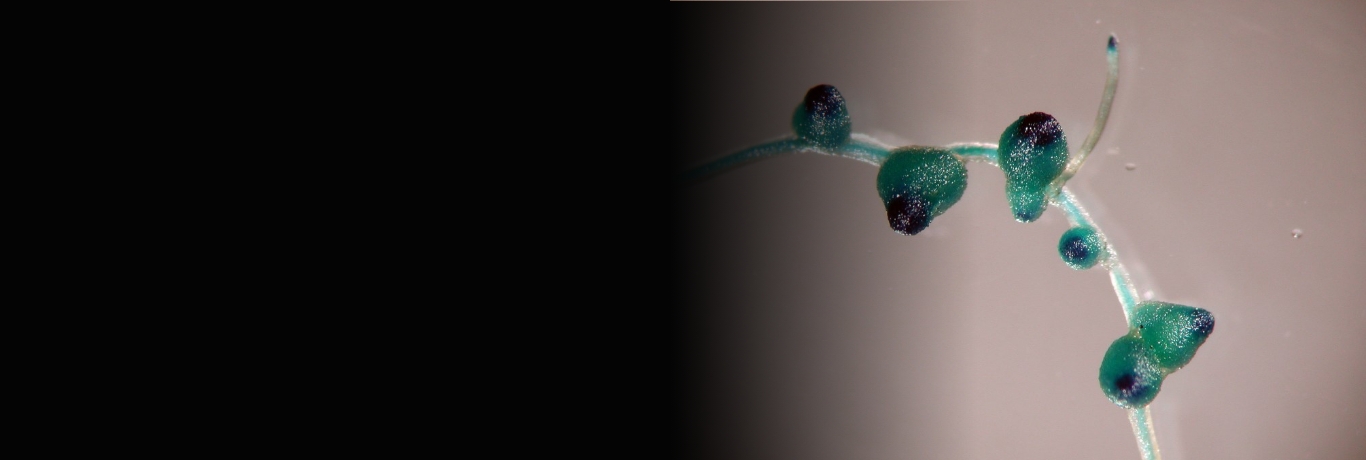
Les interactions symbiotiques, au-delà d’aboutir à la formation d’organes spécifiques (nodosités fixatrices d’azote dans le cas de l’interaction avec les rhizobia ou les actinomycètes ; ectomycorhizes) ou de cellules spécialisées (cellules corticales racinaires contenant des arbuscules dans le cas des champignons endomycorhiziens), affectent aussi le développement et le métabolisme des plantes hôtes et notamment l’architecture et les systèmes d’acquisition de nutriments de leur système racinaire.
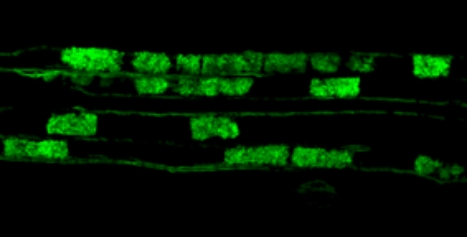
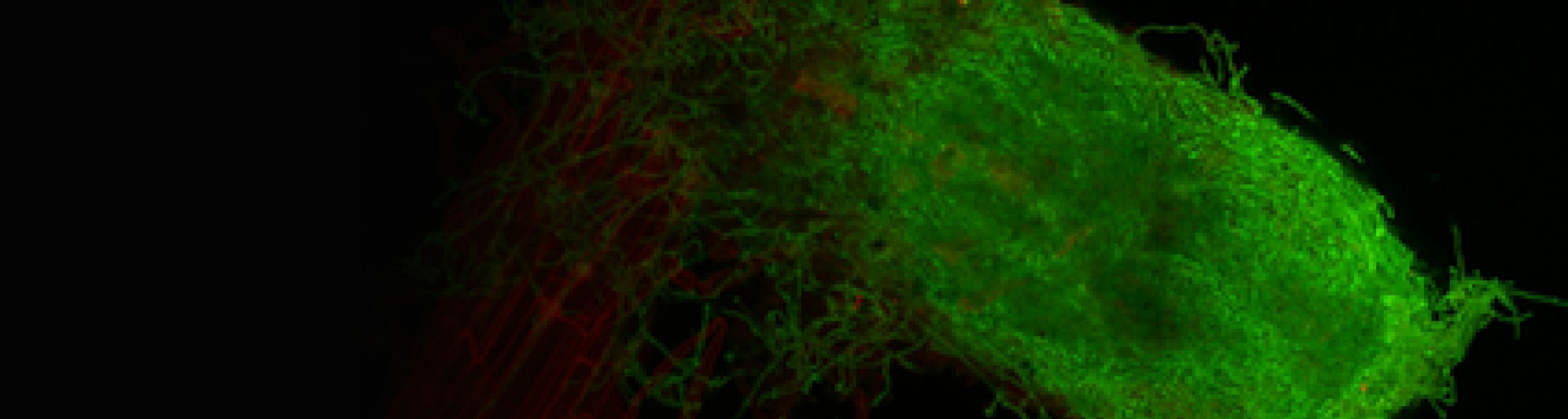
Ainsi, à la fois les rhizobia et les champignons mycorhiziens, ainsi que les signaux symbiotiques associés de type (lipo)chito-oligosaccharidiques (LCOs), promeuvent la croissance racinaire et la ramification des racines latérales. S’il a été établi que ce sont bien des réponses symbiotiques qui dépendent notamment de la voie de perception de ces signaux LCOs, les mécanismes associés, et notamment les liens avec les balances hormonales de la plante hôte restent à explorer.
De plus, le développement des nodosités symbiotiques fixatrices d’azote étant apparenté au développement des racines latérales, déterminer au niveau moléculaire quelles sont les similarités et différences entre ces deux programmes développementaux, notamment au niveau des régulations hormonales et transcriptionnelles, reste un challenge qui, s’il était relevé, permettrait d’envisager le transfert de cette capacité symbiotique à d’autres plantes que les légumineuses-hôtes.
La mise en place de ces différentes symbioses mutualistes implique aussi un contrôle développemental étroit exercé par les conditions nutritionnelles environnementales, et notamment par le niveau de disponibilité de sources minérales de nutriments tels que l’azote (N) et le phosphore (P). De plus, l’activité de ces symbioses mutualistes implique des adaptations métaboliques spécifiques associées à ces nouveaux puits de carbone / sources de nutriments. Ainsi, des régulations multiples sont déterminées par l’environnement nutritionnel minéral du sol combiné aux besoins nutritionnels de la plante-hôte, qui impliquent à la fois des signalisations au niveau local mais aussi de manière systémique au travers de l’ensemble de la plante.
La combinaison de ces régulations permet de définir à l’échelle de la plante entière les conditions permissives pour la mise en place de ces symbioses, et d’assurer une intégration entre métabolismes carbonés, azotés et phosphatés. Au-delà d’éléments moléculaires identifiés récemment (voies de perception du P et du N, rôle de peptides de signalisation…), de nombreuses questions restent à explorer pour comprendre comment les plantes-hôtes régulent l’activité symbiotique (nombre d’interactions et efficacité des échanges nutritionnels) ainsi que les adaptations métaboliques associées. Ces éléments sont tout autant essentiels à prendre en compte dans la perspective d’un transfert efficace de ces interactions symbiotiques à des plantes non-hôtes, notamment afin de ne pas générer de coût énergétique inutile lorsque les conditions environnementales ne requièrent pas la mise en place de ces symbioses.
